Das visuelle System
Physiologie des Sehens
Auge und Ohr sind Fremdempfänger (im Gegensatz z.B. zur Haut)
Gesichtsfeld Ţ was bei unbewegten Augen zu sehen ist
Blickfeld Ţ was bei unbewegtem Kopf zu sehen ist; horizontal
um 120° und vertikal 80° größer als das Blickfeld.
monoculares Gesichtsfeld
binoculares Gesichtsfeld: Sehen mit beiden Augen; große Bedeutung für dreidimensionales
Sehen
beide überlappen sich erheblich.
Perimetrie Ţ augenärztliche Ausmessung des Gesichtsfeldes. Lichtpunkte werden von außerhalb des Gesichtsfeldes auf den Fixationspunkt zu bewegt. Für weiße Lichtpunkte ist das Gesichtsfeld größer als für farbige, da das Auge in der Peripherie keine Zapfen hat, die das Farbsehen ermöglichen.
Skotome sind Gesichtsfeldausfälle auf der Netzhaut. Der blinde Fleck, der Austritt des Sehnervs, ist ein physiologisches Skotom (da in der Fovea keine Stäbchen vorhanden sind ist auch hier ein blinder Fleck bei Dämmerungssehen)
Die Sehschärfe nimmt an Deutlichkeit von der Fovea, dem Punkt des schärfsten Sehens, weg immer mehr ab. Fixationspunkt und Fovea Stimmen überein.
Photopisches und skotopisches Sehen
Das Farbensehen des Tages nennt man photopisches Sehen, das Schwarz-Weiß-Sehen der Dämmerung skotopisches Sehen. Farbensehen ist in der Dunkelheit nicht möglich. Photopisches und skotopisches Sehen werden über zwei Sensorentypen Zapfen und Stäbchen vermittelt, die in unterschiedlicher Dichte auf der Netzhaut verteilt sind. An der Fovea centralis befinden sich nur Zapfen, am Rande der Netzhaut gibt es fast nur noch Stäbchen.
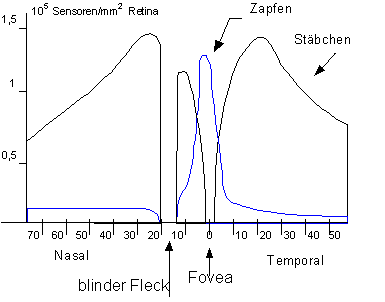
Abbildung 1: Verteilung der Stäbchen und Zapfen auf der Retina
Dunkeladaptation
Die maximale Dunkeladaptation dauert beim Menschen ca 30 Min. Zapfen adaptieren
wesentlich schneller als Stäbchen.
Helladaptation
Bei Übergang vom Dunkeln ins Helle kommt es zu einer starken Aktivierung der
Netzhautsensoren (Blendung). Die Helladaptation erreicht ihr Maximum schon nach einer
Minute.
Abbildung 2: Nachbilder
Nachbilder
Adaptiert ein Teil der Retina hell oder dunkel, und ändern sich dann die
Lichtverhältnisse, so kommt es zu negativen Nachbildern (Abbildung
2) Bei farbigen Reizmustern werden Nachbilder in der Gegenfarbe ausgelöst (rot/grün,
blau/gelb). JE intensiver die Lichtquelle, desto länger das Nachbild.
Kontrast- und Bewegungssehen
Ein physikalisch identischer Reiz wird in Abhängigkeit von seiner Umgebung
unterschiedlich wahrgenommen, so daß der Unterschied zur Umgebung betont wird (siehe
Abbildung 3). Man nennt das Simultankontrast
Die Ränder der Kreises scheinen besonders betont Dieses Phänomen wird als Mach-Bänder
bezeichnet.
Abbildung 3 : Simultankontrast

Eigengrau und Graustufen
Nach längerer Zeit im Dunkeln sehen wir nicht schwarz, sondern grau das sog.
Eigengrau. Wir erkennen die dunklen und hellen Felder eines Schachbretts auch im Dunkeln.
Fällt Licht auf das Schachbrett, so erscheinen die hellen Felder heller, die dunklen
Felder aber auch dunkler, obwohl mehr Licht von ihnen auf die Retina reflektiert wird
(Erklärung siehe Simultankontrast).
Flimmerfusionsfrequenz und Scheinbewegung
Wird eine Lichtquelle schneller als 30 mal pro Sekunde ein und ausgeschaltet, so wird
es als Dauerlicht wahrgenommen. Die FVQ hängt aber stark von der Lichtintensität ab. Je
heller das Licht, desto höher die kritischer Flimmerfrequenz. Scheinbewegungen werden
auch Phi-Phänomen genannt (Daumenkino, Lauflichter etc.)
Sehen mit zwei Augen
Durch das Sehen mit zwei Augen wird das Gesichts- und Blickfeld erweitert und es
ermöglicht uns den Abstand von Gegenständen im Raum besser abzuschätzen und ein
dreidimensionales plastisches Abbild der Umwelt aufzubauen.
Konvergenz als Entfernungsmesser
Die optischen Achsen der beiden Augen treffen sich immer im fixierten Punkt (im
Unendlichen laufen die Achsen parallel). Je näher der fixierte Punkt, desto stumpfer der
Winkel den die beiden Sehachsen bilden. Dieser Konvergenzwinkel wird vom Gehirn
festgestellt und als Maß für die Entfernung des Fixationspunktes ausgewertet.
Querdisparation und Tiefenwahrnehmung
Die seitliche Verschiebung der beiden monocularen Gesichtsfelder wird Querdisparation
genannt. Sie bewirkt, daß alle Gegenstände vor dem Fixationspunkt als gekreuzte
Doppelbilder erscheinen müßten, alle die hinter ihm liegen als ungekreuzte. Das Gehirn
gleicht diese Verschiebung zwar aus, aber die Information darüber, ob die Doppelbilder
gekreuzt oder ungekreuzt sind, werden für den räumlichen Tiefeneindruck verrechnet. Der
Tiefeneindruck wird nur bis zu einer Entfernung von sechs Metern durch die Querdisparation
gewährleistet. Bei größeren Entfernungen werden andere Informationen herangezogen (s.
u.: Größenkonstanz).
Sehen und Wahrnehmen
Um Gegenstände als solche wahrnehmen zu können bedarf es nicht nur der
Sehfähigkeit, sondern auch einiger Erfahrung und Deutung der über die Sehnerven
einströmenden Impulse.
Größenkonstanz
Ein Gegenstand wird immer, unabhängig von der Entfernung, als gleich groß angesehen,
obwohl sich die Abbildung auf der Netzhaut je nach Entfernung verändert. Das Sehsystem
verfügt über einen Mechanismus, der Größenveränderungen durch verschiedene Distanzen
ausgeglichen werden.
Formkonstanz
Bekannte Gegenstände werden immer als gleich erkannt, unabhängig von der Intensität
und der Farbe der Beleuchtung, von der Entfernung und der perspektivischen Verzerrung, vom
Blickwinkel. Wahrscheinlich treten Ergänzungs- und Kontrastprozesse auf, die über
Entfernungsmessung und Tiefenwahrnehmung (vergl. Konvergenz
und Querdisparation und Tiefenwahrnehmung) hinaus
dazu beitragen, eine geschlossene Gestalt wahrzunehmen. Das Gehirn setzt auch hier seine
Erfahrungen ein, wobei die besten Interpretation des Sinneseindruckes gewählt und als
wahr angenommen wird. Bei den Beispielen in Abbildung 5, ist keine Eindeutige Lösung des
Wahrgenommenen möglich; das Wahrnehmungssystem springt zwischen alternativen Lösungen
hin und her.
Optische Täuschungen
sind ebenfalls Fehlinterpretationen des Wahrnehmungssystems. Nicht mangelnde
Analysefähigkeit sind der Grund hierfür, sonder im Gegenteil die Fähigkeit, aus wenigen
Hinweisen eine zuverlässige Interpretation der Umwelt zu erhalten (vergl. Abbildung 6A).
Abbildung 5: unmögliche Figuren
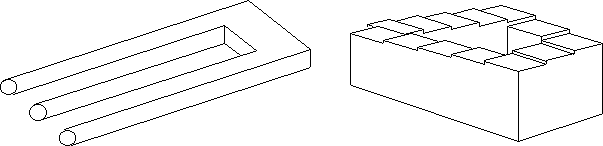
Abbildung 6: Optische Täuschungen. A) Das Rechteck existiert nicht, sondern entsteht durch Gestaltergänzung; B) Die Vertikalen sind parallel und gerade; C) Die Kreise in der Mitte sind gleich groß; D) Dir Stecken sind alle gleich lang;
Sehen, Blicken und Wahrnehmung
Wir tasten die Umgebung und Gegenstände, ‘Menschen etc. durch schnelle
Augenbewegungen, sog. Sakkaden (siehe ) ab. Konturen, Konturunterbrechungen und
-überschneidungen werden dabei häufiger angeblickt. Beim menschlichen Gesicht sind es
vor allem die Augen und der Mund; die linke Gesichtshälfte wird im Allgemeinen länger
abgetastet als die rechte.
Besonderheiten des Farbensehens
Licht und Farbe
Newton entdeckte, daß das Sonnenlicht aus allen Spektralfarben zusammengesetzt ist.
Licht ist eine elektromagnetische Strahlung mit Wellenlängen zwischen 400 und 700
Nanometern (nm). Farben haben unterschiedliche Frequenz und werden daher unterschiedlich
in einem Prisma gebrochen (chromatische Aberration). Langwelliges Licht Ţ rot, kurzwelliges Licht Ţ violett;
dazwischen liegen orange, gelb, grün, und blau (in dieser Reihenfolge). Monochromatisches
Licht Ţ wenn es nur aus einer Wellenlänge besteht.
Mischfarben können durch physiologische Mischung der Grundfarben erzeugt werden (zwei
Töne werden nicht als gemischter Ton Wahrgenommen, sondern als Akkord).
Bunte und unbunte Farben
Der farbtüchtige Mensch kann etwa 7 Millionen Farbnuancen wahrnehmen und
unterscheiden. Diese Nuancen entstehen durch die Mischung von bunten und unbunten Farben.
Die unbunten Farben setzen sich aus den Graustufen, von strahlend Weiß bis tief Schwarz,
zusammen, die bunten Farben aus dem Farbkreis siehe ). Die Sättigung einer Farbe wird
durch die den unbunten Anteil bestimmt. Die Farben braun und rosa können z.B. nur mit
unbunten Farben und nicht aus nur bunten Farben gemischt werden.
Additive und subtraktive Farbmischung
Zur additiven Farbmischung werden selbstleuchtende Farben benötigt. Jede Farbe des
Farbkreises hat eine Komplementärfarbe, die mit ihr gemischt Weiß ergibt.
Wird weißes Licht durch einen Blaufilter und einen Gelbfilter gesendet, so erhält man grünes Licht. Der Blaufilter schluckt gelbes und rotes Licht, der Gelbfilter blaues Licht. Bei Pigmentfarben (Malen) handelt es sich um subtraktive Farben da die Körner der Farben wie Farbfilter wirken.
Trichromatisches Farbensehen und die Farbtheorien
Nach Young (1773-1829) genügen drei Farben, um alle Farbtöne zu mischen. Dies drei
Primärfarben liegen nicht eindeutig fest. Man hat sich international auf die Farben mit
den Wellenlängen 700 nm (rot), 546 nm (grün) und 435 nm (blau) geeinigt (was aber
physiologisch nicht eindeutig sein muß).
Da drei Farben genügen vertritt man die Auffassung, das es auf der Netzhaut drei unterschiedlich farbempfindliche Sensorentypen gibt. Durch unterschiedlich starke Erregung dieser drei Sensoren (Zapfen) und anschließender gemeinsamer Verrechnung entstehen die unterschiedlichen Farben (Helmholtz).
Die Gegenfarbentheorie stammt von Hering und stütz sich auf das Phänomen der farbigen Nachbilder (siehe Nachbilder S.2).Demnach gibt es die vier Urfarben Rot, Gelb, Blau und Grün, wobei sich die Wirkung der Gegenfarben Rot/Grün und Blau/Gelb sowie von Schwarz/Weiß antagonistisch verhalten. Dies Erregungs- und Hemmprozesse lassen sich nicht an den Zäpfchen, aber an den nachgeschalteten Neuronen der Retina beobachten. Beide Theorien sind also auf unterschiedlichen Ebenen des visuellen Systems richtig (siehe Abbildung 7).
Farbsinnesstörungen und Farbenblindheit
Die Rot-Grün-Verwechslung ist die häufigste Farbsinnesstörung (8% Männer, 0,4%
Frauen) und beruht auf einer gestörten Empfindlichkeit des Auges für Rot oder Grün. Die
Diagnose erfolgt mit Hilfe eines Anomaloskops (ein rot-grünes Mischfeld soll durch
Veränderung der Farbkomponenten einem Gelbfeld angepaßt werden). Bei einer Rotschwäche
(Protanomale) wird mehr rot, bei einer Grünschwäche (Deuteranomale) mehr grün als
normal zugemischt. Beide sehen nicht tricromatisch sondern dichromatisch. Bei Protanopen
ist das Farbspektrum am langwelligen Ende verkürzt (rotblind); für den Deuteranopen gilt
Entsprechendes.
Bei Gelb-Blau-Verwechslung ist das blau-violette Ende des Spektrums verkürzt; kommt
sehr selten vor.
Die totale Farbenblindheit heißt Achromasie oder Monochromasie und kommt durch den
völligen Ausfall der Zapfenfunktion zustande. Es ist nur ein skotopisches Sehen möglich
und am Tage müssen Sonnenbrillen getragen werden; die Sehschärfe ist um 10% reduziert.
Die Untersuchung des Farbsinns erfolgt über pseudo-isochromatische Tafeln (Sehtest).
Das Auge
Das optische System des Auges
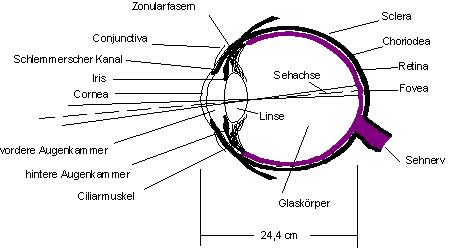
Das "Objektiv" des Auges ist zusammengesetzt aus der Cornea (Hornhaut), vorderer Augenkammer, und Linse. Es entwirft auf der Netzhaut ein umgekehrtes und stark verkleinertes Bild. Die Sehachse trifft auf die Zentralgrube (Fovea centralis oder gelber Fleck), die Stelle des schärfsten Sehens.
Die Iris dient als Blende, die bei hellen Lichtquellen die Pupille mit Hilfe des Ciliarmuskels? verkleinert und sie im Dunkeln weitet. Die Iris beider Augen sind synchron geschaltet.
Zur Scharfeinstellung von Gegenständen in unterschiedlichen Entfernungen wird die Linse des Auges mehr oder weniger (bei nahen Gegenständen stark, bei weit entfernten kaum bis gar nicht) gekrümmt und verändert somit ihre Brechkraft.
Die Linse des Auges ist an sog. Zonularfasern befestigt, die in den Ciliarmuskel münden. Durch Kontraktion des Ciliarmuskels wird die Linse gekrümmt. Bei erschlafften Muskel sorgen die Zonularfasern und der Innendruck des Auges dafür, daß die Linse wieder abflacht.
Die lichtempfindliche Schicht des Augenhintergrundes ist die Netzhaut (Retina). Sie besitzt zwei Typen von Photosensoren, die Stäbchen und Zapfen, sowie ein Netzwerk nachgeschalteter Nervenzellen (die letzte Schicht sind die Ganglienzellen). Die Ganglienzellen sammeln sich zu einem Bündel, dem Sehnerv, der etwa auf der Höhe der Fovea das Auge verläßt. Die Netzhaut ist so aufgebaut, daß die Lichtreize zunächst durch die gesamte Neuronenschicht laufen, bevor sie auf die Photosensoren treffen, was zu Leistungseinbußen im Vergleich zu Kameras führt. Nur in der Fovea liegen die Zapfen frei, so daß Lichtreize diese unmittelbar treffen. Die Zapfen in der Fovea sind besonders gut mit dem Gehirn verschaltet, während sich die übrigen Sensoren auf der Retina Neurone teilen.
Abbildung 7: Der Aufbau des Auges
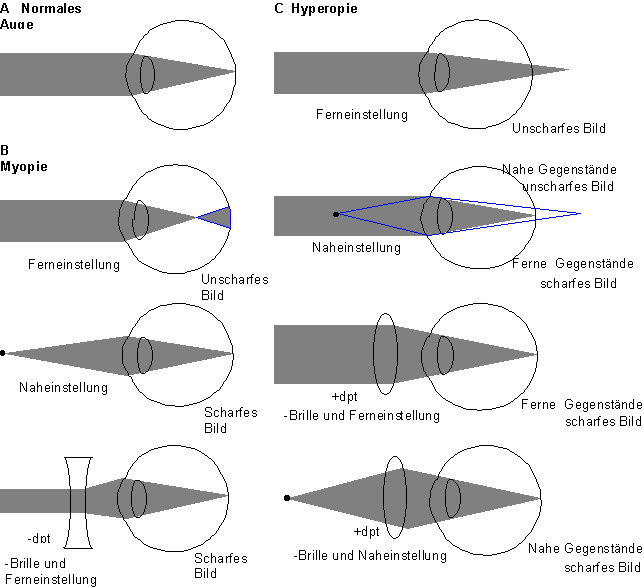
Brillen für nah und fern
Ist die Entfernung von der Cornea bis zur Oberfläche der Retina größer oder kleiner als
24,4 mm, so kommt es zur Kurz- bzw. Weitsichtigkeit (vergl. Abbildung 8) Die Lichtstrahlen
eines Gegenstandes treffen sich dann nicht in der Fovea, sondern davor oder dahinter.
Kurzsichtigkeit (Myopie). Der Augapfel ist zu lang und die Strahlen treffen vor der Retina zusammen. Die Brechkraft ist relativ zur Augapfellänge zu groß. Nahe Gegenstände werden scharf gesehen. Myopie wird durch eine Zerstreuungslinse kompensiert. Die Stärke der Linsen wird in Dioptrie angegeben und berechnet sich durch den Kehrwert der Brennweite.
Weitsichtigkeit (Hyperonie). Ist der Augapfel zu kurz, treffen sich die aus dem unendlichen kommenden Strahlen hinter der Retina. Durch Naheinstellen (Erhöhung der Brechkraft der Linse) können entfernte Gegenstände scharf gesehen werden. Für die Scharfeinstellung naher Gegenstände reicht die Brechkraft nicht mehr aus. Kompensation dieser Schwäche erfolgt durch Sammellinsen.
Alterssichtigkeit (Presbyopie). Im Alter verliert die Liste ihre Elastizität und eine Scharfeinstellung in der Nähe ist nicht mehr möglich.
Beim Astigmatismus ist die Cornea in einer Richtung stärker gekrümmt als in der anderen. Bei Differenzen > 0,5 dpt wird der Astigmatismus durch Gläser ausgeglichen werden, die nur in derjenigen Achse geschliffen sind, die auf der Cornea zu stark gekrümmt ist.
Abbildung 8: Strahlengang bei normalen und fehlsichtigen Augen.
Die Signalverarbeitung im visuellen System
Durch die Weiterleitung der Sinnesreize wird kein Bild im Gehirn abgebildet, sondern in Impulsmuster verschlüsselte Informationen, also ein Abbild der vom Auge betrachteten Objekte.
Die Signalverarbeitung in der Netzhaut
Transduktion in den Photosensoren.
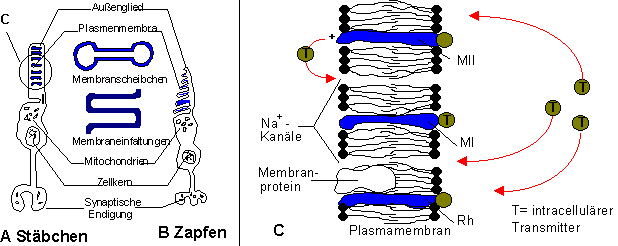
Auf der Retina befinden sich etwa 20 Millionen Stäbchen und 6 Millionen Zapfen in
unterschiedlicher Verteilung (siehe oben). Das obere Ende der Sensoren besteht aus ca
tausend Membranscheibchen (Stäbchen) bzw. -einfaltungen(Zapfen) in denen der Sehfarbstoff
eingelagert ist (Abbildung 9A,B). Über die Gewebsbrücke (Cilium) sind Zapfen und
Stäbchen mit dem Zellkörper verbunden, der synaptisch mit den nachfolgenden Neuronen
verbunden ist. Licht wird von den Sehfarbstoffen absorbiert und leitet durch den dadurch
bewirkten Zerfall in Vorstufen das hyperpolarisierende Sensorpotential (bei
anderen Sensoren depolarisierend) ein. Der Sehfarbstoff der Stäbchen heißt Rhodopsin
(Sehpurpur). Er zerfällt in das Eiweiß Opsin und Vitamin A, aus denen er durch
Energieaufwand wieder aufgebaut wird (Abbildung 9C, S.*). In großer Helligkeit ist der Farbstoff nahezu ausgebleicht, bei
Dunkelheit ist er bis zum Max. regeneriert. Je mehr Sehpurpur vorhanden ist, desto
größer die Lichtempfindlichkeit. Mangel an Vitamin A, das zum Aufbau des Farbstoffs
nötig ist, führt zu Nachtblindheit.
Die chemische Struktur der Zapfen ist nicht bekannt. Es existieren drei Typen, die das trichromatische (Farben)sehen ermöglichen (vergl. oben).
Abbildung 9: Transduktion in der Retina
Signalverarbeitung in den retinalen Ganglienzellen
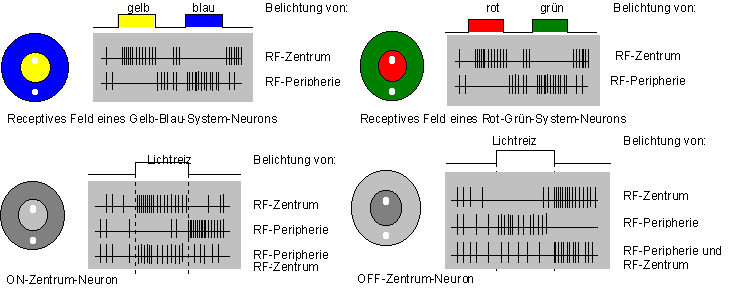
Zwischen den Photosensoren und den Ganglienzellen liegen die retinalen Neurone. Zwei Hauptflußrichtungen der Information sind zu unterscheiden: Zentripedal über die Bipolarzellen und quer dazu über Horizontalzellen und Amakrinen ebenfalls zu den Ganglienzellen. Die neuronale Reizverarbeitung verläuft ausschließlich über langsame lokale Membranpotentiale. 125 Millionen Photosensoren teilen sich dabei nur 1 Millionen Ganglienzellen. In den Ganglienzellen findet schon eine erheblicher Teil der Informationsverarbeitung statt. Ihre Axone verlassen im Sehnerv das Auge und sie bilden im Gegensatz zu den vorgeschalteten Zellen Aktionspotentiale aus. Fällt ein Lichtreiz auf eine Stelle der Retina, so ändert sich die Aktivität der Ganglienzellen. Das Zentrum eines so gereizten "receptiven Feld" verhält sich antagonistisch zu seiner Peripherie. Wirkt ein Reiz auf das Zentrum erregend so hat das hemmende Wirkung auf die Peripherie. Es werden zwei Arten von Zentren unterschieden: die On- und Off-Zentren. Die On-Zentren entladen sich während eines Reizes, die Off-Zentren nach Beendigung dieses Reizes (vergl. Abbildung 10 S.*). Sie stellen die wichtigsten Klassen der retinalen Ganglienzellen dar. Die receptiven Felder sind in der Fovea centralis am kleinsten und vergrößern sich kontinuierlich in Richtung Peripherie, das heißt, in der Fovea ist das Auflösungsvermögen am größten. Die Größe von Zentrum und Peripherie ist variabel. Bei Helladaptation ist das Zentrum klein und die Peripherie groß. Bei Dunkelheit verhält es sich umgekehrt oder die Peripherie ist gar nicht mehr nachzuweisen.
Die Verarbeitung farbiger Lichtreize in retinalen Ganglienzellen folgt im Allgemeinen denselben Gesetzmäßigkeiten wie oben besprochenen. Statt des Hell-Dunkel-Antagonismus sprechen wir hier von Gelb-Blau- und Rot-Grün-Antagonismus. Wird ein "rotes" Stäbchen gereizt, wirkt das auf die grüne Peripherie hemmend und umgekehrt. Für die Gelb-Blau-System-Neurone gilt entsprechendes.
Abbildung 10: Funktionelle Organisation receptiver Felder
Die Verarbeitung in subcorticalen visuellen Zentren
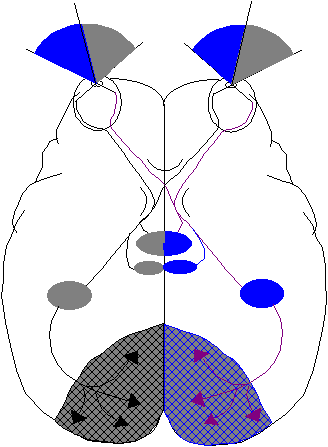
Die Sehnerven (nervi optici) beider Augen tauschen in der Sehkreuzung, dem Chiasma opticum, etwa die Hälfte ihrer Nervenfasern aus, und zwar so, daß die linke Gesichtshälfte beider Augen zur rechten Gehirnhälfte projiziert werden und umgekehrt (vergl. Abbildung 11, S.*). Nach dem Chiasma opticus laufen die Ganglienaxone zu einem Thalamuskern, dem Corpus geniculatum laterale (CGL) oder seitlicher Kniehöcker. Hier findet die einzige synaptisch Umschaltung statt. Die sog. Sehstrahlung (Radiato optica) führt von hier aus zur Sehrinde (visueller Cortex) im Hinterhauptlappen (Areal 17 nach Brodman). Es bestehen zahlreiche Verbindungen zu anderen Arealen, (insb. 18 u.19). Diese Zentren sind über den Balken (corpus callosum) mit der anderen Hirnhälfte verbunden.
Weitere Kollaterale entspringen unmittelbar nach dem Chiasma und führen zum Hirnstamm, in Kerne (vordere vier Hügel oder Colliculi superiores), die für die Augenmotorik zuständig sind. Die Neurone dieser Kollateralen reagieren besonders auf bewegte Reizmuster.
Das CGL besteht aus 6 Neuronenschichten, in denen die Informationen der beiden Augen (noch) getrennt verarbeitet werden. In den Schichten 2,3 und 5 finden sich Neurone des ipsilateralen, in 1,4 und 6 die des kontralateralen Auges. Erst im Visuellen Cortex wird also die Information beider Augen binocular, zum Zwecke des stereoskopischen Sehens verarbeitet.
Die Signalverarbeitung im visuellen Cortex
Die topologische Organisation der Sehrinde erstreckt sich über den Gyrus postcentralis. Das Gebiet um die Fovea nimmt hier etwa die Hälfte des Areals ein. Die Nervenfasern der Sehstrahlung enden Schicht IV des Cortex. Die Verarbeitung erfolgt in sog. corticalen Säulen von Neuronen (senkrecht angeordnet). Säulen mit Neuronen des rechten wechseln sich mit denen des linken Auges ab. Sie werden oculäre Dominanzsäulen genannt. Eine weitere Unterteilung in Unter- bzw. Orientierungssäulen trennt solche, die durch verschiedene Qualitäten des Sehens (Farbe, Orientierung von Strukturen, bewegte Reize nach Richtung getrennt etc.) erregbar sind.
Komplexere receptive Felser kon´mmen in den höheren Schichten des Cortex (III,II,I) vor. Sie reagieren auf bewegte Reizmuster stärker als auf unbewegte.
Abbildung 11: Schema der Sehbahnen im Gehirn des Menschen